
潛水鳥屬
| 潛水鳥屬 化石時期:晚白堊世,
| |
|---|---|

| |
| 繪於1880年的跗跖骨素描圖 | |
| 科學分類 | |
| 界: | 動物界 Animalia |
| 門: | 脊索動物門 Chordata |
| 綱: | 蜥形綱 Sauropsida |
| 總目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亞目: | 獸腳亞目 Theropoda |
| 演化支: | 鳥翼類 Avialae |
| 科: | †潛水鳥科 Baptornithidae AOU, 1910 |
| 屬: | †潛水鳥屬 Baptornis Marsh, 1877 |
| 模式種 | |
| †外來潛水鳥 Baptornis advenus Marsh, 1877[1]
| |
| 異名 | |
|
副斯堪尼鳥 Parascaniornis | |
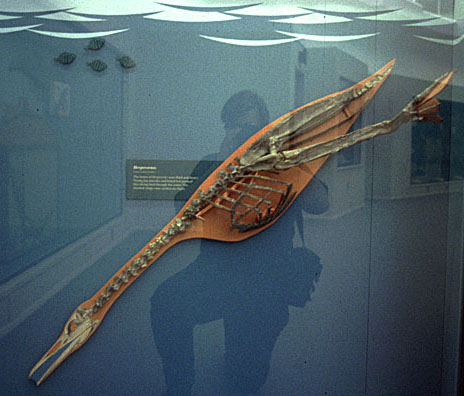
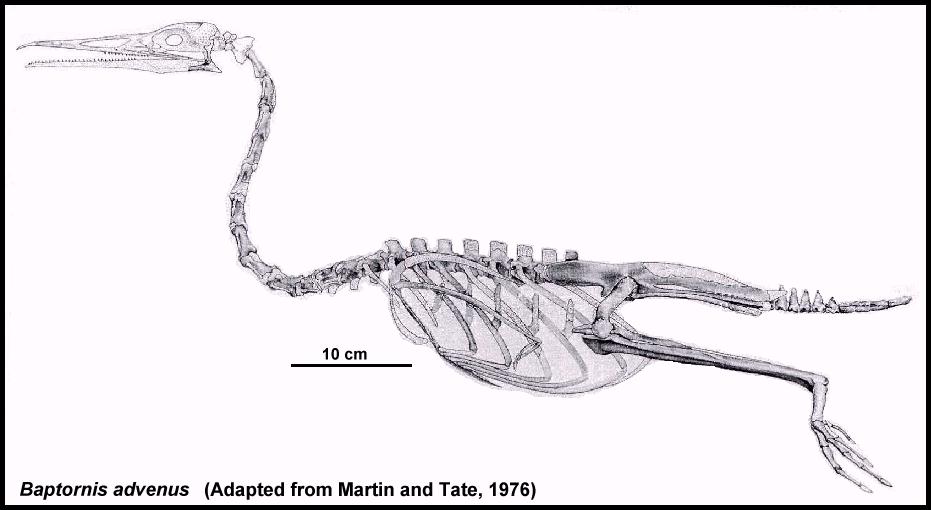
潛水鳥屬(學名:Baptornis)是種不飛會的史前水鳥,生存於大約8700至8000萬年前的晚白堊世(大致為中科尼亞克期至中坎帕期)。模式種是外來潛水鳥(Baptornis advenus),化石發現於堪薩斯州,當時為稱作西部內陸海道的淺陸架海所覆蓋。潛水鳥亦分布於白堊紀的瑞典,當時圖爾蓋海峽尚與古代北海相連接。該屬的存在可能遍及整個全北界。
首個化石由奧塞內爾·查利斯·馬什於19世紀70年代發現,和始祖鳥一樣都是最早為學界所知的中生代鳥類之一。
生境
[編輯]關於外來潛水鳥的生境,可能存在較其它黃昏鳥類更多的材料證據(帝王黃昏鳥可能除外),但仍有許多有待研究。這隻潛鳥大小的鳥,在其近親中屬中等體型,有着異常修長的頸部。推測其行為類似今天的蛇鵜,獵取比其大型親戚更小更靈活的獵物。但與蛇鵜不同的是潛水鳥無法用嘴刺穿獵物,而是像今代的秋沙鴨一樣用喙捉取獵物。
潛水鳥所棲息的水域為相當淺的陸緣海或陸架海。在遠離史前海岸區域發現的遺骸,表明其很可能冒險到很遠的地方並(或)在島嶼上繁殖。已發現相當數量的幼鳥標本。這些標本一般出土於該屬分布範圍的北部,即如今的加拿大和阿拉斯加,儘管也有一些在堪薩斯發現。這表明潛水鳥可能像現在的企鵝一樣遷徙,在夏季前往極地繁殖。白堊紀的氣候較今天溫暖許多,潛水鳥棲息的水域為亞熱帶至溫帶氣候。
潛水鳥善於游泳和潛水,但人們認為其在陸地上非常笨拙,用腳推動自己沿岩石滑動而非真正地行走。小腿自然位置與身體齊平,雙腳側向伸展,故不會出現不趴下就無法豎直移動的情況。與只能像海豹一樣用腹部滑行的黃昏鳥相反,潛水鳥小腿的活動泛圍並非只局限於身體兩側,因此更容易把腳放在身體下面,同時腳趾指向前方,行走時可能有小步跳躍甚至笨拙的搖擺,身體則低垂於地面。
關於黃昏鳥目的食物,目前唯一明確記錄來自潛水鳥:標本UNSM 20030中發現了糞化石,為直徑一厘米左右的圓形小塊,內含「劍齒鯡魚」矛齒魚的殘骸,具體物種可能為小矛齒魚。潛水鳥有強大的胃液,且(或)能像大多數現存食魚鳥類一樣,反芻獵物大多數難以消化的部分形成食丸,因為矛齒魚殘骸僅占糞化石的一小部分,大部分則為普通的糞便。
分類
[編輯]潛水鳥與體型更大的黃昏鳥近緣,二者均屬黃昏鳥目,這是一群適應潛水和游泳的獨特有齒鳥類[2]。與始祖鳥或反鳥類不同,它們的外觀更類似現存鳥類而非恐龍。
本屬在黃昏鳥目中非常獨特,故為其建立單獨的潛水鳥科(Baptornithidae),有很多人都認為它是單系群。但近期研究提出,生存於晚白堊世的瑞典伊沃島、人稱「白堊紀火烈鳥」的斯氏副斯堪尼鳥(Parascaniornis stensioi)並非火烈鳥,也不像其他人提出的那樣屬於潛鳥目或鸌形目,實際上應屬於潛水鳥。但由於化石稀少無法對比,目前尚不清楚它是外來潛水鳥的次異名還是該屬的第二個種[3]。
2004年宣布正在準備第二件材料以供描述,該標本重量約為外來潛水鳥正模的兩倍。這些骨頭發現於南達科他州西南的皮埃爾頁岩下段[4]。詹姆斯·馬丁(James Martin)與阿曼達·科爾斯-珀森(Amanda Cordes-Person)於2007年將此物種命名為瓦氏潛水鳥(Baptornis varneri),但後來將其重新歸入可能與外來種近緣的布氏鳥屬[5]。
此外,還有兩種晚白堊世的史前潛鳥有時也被歸入潛水鳥科。河流鳥很可能是黃昏鳥目成員,但目前還不清楚它與黃昏鳥目的哪一科關係最近,有些研究將其歸入潛水鳥科。更有爭議的是新潛鳥。這種鳥類的遺骸發現於智利,可能也屬於潛水鳥科。另一些人則認為其與某些現代鳥類如潛鳥目或鸌形目近緣。
譜系關係
[編輯]2015年,一項物種水平的系統發育分析發現黃昏鳥目之間存在以下關係[6]。
| 黃昏鳥目 Hesperornithes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
參考資料
[編輯]- ^ Brands, Sheila. Taxon: Genus †Enaliornis. Project: The Taxonomicon. 7 Apr 2012 [12 Jun 2012]. (原始內容存檔於2014-02-21).
- ^ Everhart, Mike. Hesperornis regalis Marsh 1872 - Toothed marine birds of the Late Cretaceous seas. Oceans of Kansas. 2006 [23 Aug 2007]. (原始內容存檔於1999-10-06).
- ^ Rees, Jan; Lindgren, Johan. Aquatic birds from the Upper Cretaceous (Lower Campanian) of Sweden and the biology and distribution of hesperornithiforms.. Palaeontology. 2005, 48 (6): 1321–1329. Bibcode:2005Palgy..48.1321R. doi:10.1111/j.1475-4983.2005.00507.x
 .
.
- ^ Person, Amanda Cordes. A New Species of Diving Bird, Baptornis, from the Lower Pierre Shale (Upper Cretaceous) of Southwestern South Dakota.. Rocky Mountain (56th Annual) and Cordilleran (100th Annual) Joint Meeting: 33–37. May 2004.
- ^ Martin, Larry D.; Evgeny, Kurochkin; Tocaryk, Tim T. A new evolutionary lineage of diving birds from the Late Cretaceous of North America and Asia. Palaeoworld. 2012, 21: 59–63. doi:10.1016/j.palwor.2012.02.005.
- ^ Bell, A.; Chiappe, L. M. A species-level phylogeny of the Cretaceous Hesperornithiformes (Aves: Ornithuromorpha): Implications for body size evolution amongst the earliest diving birds. Journal of Systematic Palaeontology. 2015, 14 (3): 239–251. S2CID 83686657. doi:10.1080/14772019.2015.1036141.
外部連結
[編輯]- Mounted skeleton. Lobed feet less likely, but plausible. Retrieved 2007-AUG-23.
- Reconstructed skeleton. Retrieved 2007-AUG-23.
- Reconstruction in life. Color is based on reasonable assumption of countershading. Retrieved 2007-AUG-23.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







.png)















{kind=link}
{kind=link}
{kind=link}